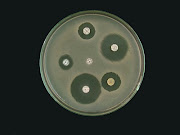
Objectives:
1. To produce and extract protease from Bacillus spp.
2. To study of its enzymatic activity and compare with standard
Introduction:
ENZYME
All living organism depend on catalytic proteins called enzymes, these compound are responsible for metabolism and biochemical regulation of energy, there are millions of enzymes and hundreds of enzymes have been identified in the human body as well. All living system possesses enzymes as vital components of the body, enzymes are found in all organisms. Life is based on the interaction of enzymes within the body of an organism, thus they are critical for life and necessary in all living systems and near living replicating systems like viruses. The identification to date of three thousand different enzymes in the human body by scientists is an indication of the myriad roles that enzymes play in the living body. Enzymes are degraded and renewed almost every second in the human body mostly at an unbelievable rate. They perform numerous chemical functions vital to the existence and survival of the body.
Enzymes have a great specificity, which means they are very specific on type of substrate they act on. Usually, every single enzyme in the body will catalyze and promote only one particular type of chemical reaction - this is often strictly so, thus they are very specific on what they act on. Different enzymes carry out different reactions in the body, thus some types of enzymes help in the degradation of large nutrient compounds in food into smaller molecules for digestion and ultimate absorption at the cellular level - these are proteins, carbohydrates, and lipids or fats. Some types of enzymes are solely involved in distinct functions, including the storage and release of biochemical energy. Other enzymes catalyze the cellular processes of respiration, other enzymes are involved in the process of reproduction, some are involved in vision and sensory reactions, etc.
Classification of enzymes
There are many classification schemes for enzymes, one example, is to classify them according to the substrate they act on, thus proteases act on proteins, lipases of lipids and amylases on carbohydrates. In the most common scheme, all enzymes are classified with respect nature of the reaction they catalyze. In this scheme, enzymes are divided into six main groups, each group possessing fundamentally different activities and catalyzing a unique reaction. The enzymes classified into the six groups according to the reactions induced are 1) oxidoreductases 2) transferases 3) hydrolases 4) lyases 5) isomerases, and 6) ligases. A brief discussion on these reactions follows below.
1. Oxidoreductases
are an important class of enzymes that have a pivotal role in metabolism inside the cell. As the name suggests, this class of enzyme is responsible for oxidation and reduction processes that are always coupled in the cell. There are many ways to define oxygenation or oxidation, one way would be to speak of the addition of one oxygen molecule or removing two hydrogen atoms from a molecule, the process of reduction is just the opposite and occurs concurrently. During the oxidation process, one substrate is oxidized and another is always reduced, thus these two processes are linked and simultaneous.
2. Transferases
this class of enzymes is responsible for the catalysis of biochemical reactions during which the ions other than hydrogen are transferred between the reactants. A good example of this type of enzyme is the class known as the transamidases that transfer amide ions across reactants; many medical diagnostic techniques employ these enzymes.
3. Hydrolases
this class of enzymes are involved in the catalysis of chemical reactions in which the chemical bonds are broken off through the addition of a molecule of water. More properly, it can be understood as the addition of hydrogen [H] to a fragment of the molecule while a hydroxide [OH] ion is fixed to the other molecule fragment. This important class of enzymes includes the familiar proteolytic protein digesting enzymes, the lipolytic fat digesting enzymes, and the amylolytic carbohydrate digesting enzymes.
4. Lyases
this class of enzymes is responsible for catalyzing reactions that result in the rearrangement of the molecular structures within the reactants. This action results in the chemical conversion of a molecule into another related molecule called an isomer of the original molecule - the process is called isomerization. The chemical conversion of a D-form of a sugar to an L-form of a sugar would be a good example of an isomerization reaction catalyzed by such enzymes.
5. Isomerases
are enzymes that catalyze only isomerization reactions. These enzymes are responsible for the chemical interconversion of aldose to ketose sugars, as an example.
6. Ligases
are enzymes that actively catalyze the formation of a chemical bond between two unrelated molecules. They add molecular groups to a substrate.
Proteases
- Introduction
Proteolytic enzymes or proteases catalyse the cleavage of peptide bonds in proteins. They are enzymes of class 3, the hydrolases, and subclass 3.4, the peptide hydrolases or peptidases. They constitute a large family (EC 3.4) divided as endopeptidases or proteinases (EC 3.4 21-99) and exopeptidases (EC 3.4.11-19) according to the point at which they break the peptide chain. These endopeptidases can be ordered further, according to the reactive groups at the active site involved in catalysis, into serine- (EC 3.4.21), cysteine- (EC 3.4.22), aspartic-proteinases or-endopeptidases (EC.3.4.23) and metalloproteinases or metalloendopeptidases ( EC 3.4 24 ). Enzymes whose reaction mechanism has not been completely elucidated are classed in the subgroup EC. 3.4. 99.
- Exopeptidases
The exopeptidases act only near the ends of polypeptide chains at the N or C terminus. Those acting at a free N terminus liberate a single amino acid residue (aminopeptidases), a dipetide (dipeptidyl- peptidases) or a tripeptide (tripeptidyl- peptidases). The exopeptidases acting at a free C terminus liberate a single amino acid (carboxypeptidases) or a dipeptide (peptidyl-dipeptidases). Some exopeptidases are specific for dipeptides ( dipeptidases ) or remove terminal residues that are substituted, cyclized or linked by isopeptide bonds. Isopeptide bonds are peptide linkages other than those of a carboxyl to a -amino groups and this group of enzymes is re denominated omega peptidases.
- Endopeptidases
Endopeptidases act preferentially in the inner regions of peptide chains away from the N and C termini. The presence of free a-amino or a- carboxyl groups has a negative effect on the activity of the enzyme.
- Catalytic site
Hartley in 1960 described four distinct types of catalytic mechanisms used by peptidases. This observation has been developed into a practical system of classification (Barret,1994) . In this classification, the carboxypeptidases and the endopeptidases are divided into sub-classes on the basis of catalytic mechanism. The carboxypeptidases are subdivided in serine-, metallo and cysteine- type carboxypeptidases and the endopeptidases are the serine-, cysteine-, aspartic- and metalloendopeptidases. Serine peptidases have a serine residue involved in the active center, the aspartic have two aspartic acid in catalytic center, cysteine -type peptidases have a cysteine residue and metallo-peptidases use a metal ion in the catalytic mechanism.
Classification of Protease
Standard
Proteases are currently[update] classified into six broad groups:
- Serine proteases
- Threonine proteases
- Cysteine proteases
- Aspartate proteases
- Metalloproteases
- Glutamic acid proteases
The threonine and glutamic-acid proteases were not described until 1995 and 2004, respectively. The mechanism used to cleave a peptide bond involves making an amino acid residue that has the cysteine and threonine (proteases) or a water molecule (aspartic acid, metallo- and glutamic acid proteases) nucleophilic so that it can attack the peptide carboxyl group. One way to make a nucleophile is by a catalytic triad, where a histidine residue is used to activate serine, cysteine, or threonine as a nucleophile.
Within each of the broad groups proteases have been classified, by Rawlings and Barrett, into families of related proteases. For example within the Serine proteases families are labelled Sx where S denotes the serine catalytic type and the x denotes the number of the family, for example S1 (chymotrypsins). An up to date classification of proteases into families is found in the MEROPS database.
By optimal pH
Alternatively, proteases may be classified by the optimal pH in which they are active:
- Acid proteases
- Neutral proteases, notably involved in type 1 hypersensitivity. Here, it is released by mast cells and causes activation of complement and kinins.[3] This group includes the calpains.
- Basic proteases (or alkaline proteases)
Proteases: function and application
Proteases represent a class of enzymes with important roles in physiological process. Besides this, they have commercial applications, being one of the three largest group of industrial enzymes, accounting for about 60% of the total worldwide sale of enzymes. These enzymes are involved in essential biological processes like blood clotting, controlled cell death, and tissue differentiation. They catalyse important proteolytic steps in tumor invasion or in infection cycle of a number of pathogenic microorganisms and viruses. This makes proteases a valuable target for new pharmaceuticals. They also participate in protein catabolism in degradative or biosynthetic pathways and in the release of hormones and pharmacologically active peptides from percursor proteins. They conduct highly specific and selective modifications of proteins such as activation of enzymes by limited proteolysis and collaborate with the transport of secretory proteins across membranes. Besides this they have a variety of applications, mainly in the detergent and food industries. In view of the recent trend of developing environmentally friendly technologies, proteases are envisaged to have extensive applications in leather treatment and in several bioremediation processes, and in pharmaceutical industry for preparation of medicines such ointments for debridement of wounds. Proteases assist the hydrolysis of large polypeptides in smaller peptides and aminoacids, thus facilitating their absorption by the cells. The extracellular enzymes play a major hole in nutrition due to their depolimerizing activity.
Microbial proteases
Proteinases are found in several microrganisms such as viruses, protozoa, bacteria, yeast and fungi. The inability of the plant and animal proteases to meet current world demands has led to an increased interest in microbial proteases. Microorganisms represent an excellent source of enzymes owing to their broad biochemical diversity and their susceptibility to genetic manipulation. Proteins are degraded by microorganisms, and they utilize the degradation products as nutrients for their growth. The degradation is initiated by proteinases (endopeptidases) secreted by microorganisms followed by further hydrolysis by peptidases (exopeptidases) at the extra- or intracellular site. Numerous proteinases are produced by microorganisms depending on the species of the producers or the strains even belonging to the same species. Several proteinases are also produced by the same strain under various cultural conditions. Candida albicans and C. tropicalis are the medically more important opportunistic pathogens causing infections in immunocompromised patients. Their extracellular enzyme, an aspartic proteinase is considered to be a major virulence factor. Human immunodeficiency virus (HIV) a causative of AIDS, presents an aspartic protease essential for the retroviral life cycle and it has been a good target for the chemotherapy with specific inhibitors. Most commercial serine proteases, mainly neutral and alkaline, are produced by organisms belonging to the genus Bacillus. Representatives of this group are subtilisins. Similar enzymes are also produced by other bacteria such as Thermus caldophilus and Desulfurococus mucosus, Streptomyces , Aeromonas and Escherichia genera. Fungi produce several serine proteinases. Among them, these enzymes are produced by various strains of Aspergillus oryzae. Cysteine proteinases are not so widely distributed as was seen with serine and aspartic proteinases. However in protozoa belonging to the Trypanosomatidae family they are found as well as metaloproteinases, in several genus such as Crithidia, Phytomonas, Herpetomonas, Trypanosoma, Leishmania , Leptomonas and Endotrypanum are involved with the nutrition, life cycle and morphological differentiation of these parasites. Trichomonas vaginalis, a flagellated protozoan responsible for trichomonosis, one of the most common sexually transmitted diseases, has numerous cysteine- and some metallo-proteinases. The cysteine enzymes are involved in the damage to the host by the parasite. Sufficient data have been gathered to demonstrate unequivocally that proteases are ubiquitous in microrganisms. Microbial proteases account to approximately 40% of the total worldwide enzymes sale. In addition, proteases from microbial sources are preferred to the enzymes from plant and animal souces since they possess almost all characteristics desired for their biotechnological applications.
Reagents and Preparation
1. Protease (crude and standard)
2. Phosphate Buffer pH-7.00
3. Casein, 1 % in PB Buffer
4. Trichloroacetic Acid, 5 %
5. Folin & Ciocalteu’s Phenol Reagent
6. Alkaline Sodium Carbonate solution
7. Acetate Buffer
8. L-Tyrosine, Free Base
Before beginning the assay, we need to make sure that the following reagents are correctly prepared:
- A 50 mM Potassium Phosphate Buffer, pH 7.5. Prepare using 11.4 mg/ml of potassium phospate dibasic, trihydrate in purified water and adjusting pH with 1M HCl. This solution is placed at 37°C prior to use.
- A 0.65% weight/volume casein solution, prepared by mixing 6.5 mg/ml of the 50 mM potassium phosphate buffer. The solution temperature is gradually increased with gentle stirring to 80-85 °C for about 10 minutes until a homogenous dispersion is achieved. It is very important not to boil the solution. The pH is then adjusted if necessary with NaOH and HCl.
- A 110 mM Trichloroacetic acid solution, prepared by diluting a 6.1N stock 1:55 with purified water. Trichloroacetic acid is a strong acid and should be handled with care.
- 0.5 M Folin & Ciolcaltea’s, or Folin’s Phenol Reagent, which is the solution that will react with tyrosine to generate a measurable color change that will be directly related to the activity of proteases. Folin’s Phenol Reagent is an acid and should be handled with care.
- A 500 mM Sodium Carbonate solution, prepared using 53 mg/ml of anyhydrous sodium carbonate in purified water.
- An enzyme diluent solution, which consists of 10 mM Sodium Acetate Buffer with 5mM Calcium, pH 7.5, at 37°C. This solution is what we use to dissolve solid protease samples or dilute enzyme solutions.
- 1.1 mM L-tyrosine Standard stock solution. Prepared using 0.2 mg/ml L-tyrosine in purified water and heated gently until the tyrosine dissolves. As with the casein, do not boil this solution. Allow the L-tyrosine standard to cool to room temperature. This solution will be diluted further to make our standard curve.
If necessary, a solid protease sample of predetermined activity, which is dissolved using enzyme diluent to 0.1-0.2 units/ml. This solution serves as a positive control for the quality control assay and as validation for the calculations we will perform to determine enzyme activity.
Procedure:
Setting up the Protease Assay and Standard Curves
To begin this assay, find suitable vials that will hold about 15 mls. For each enzyme that you will test, you will need 4 vials. One vial will be used as a blank, and three others will be used to assay activity of three dilutions of the protease. Three dilutions are useful when checking our final calculations against each other. To each set of four vials add 5mls of our 0.65% casein solution, and let them equilibrate in a water bath at 37°C for about 5 minutes. Then, add varying volumes of enzyme solution you want to test to three of the test sample vials, but not the blank. Mix them by swirling and incubate for 37°C for exactly ten minutes. The protease activity and consequential liberation of tyrosine during this incubation time is what will be measured and compared between our test samples.
After this 10 minute incubation, add the 5 mls of the TCA reagent to each tube to stop the reaction. Then an appropriate volume of enzyme solution is added to each tube, even the blank, so that the final volume of enzyme solution in each tube is 1 ml. This is done to account for the absorbance value of the enzyme itself and ensure that the final volume in each tube is equal. Now incubate the solutions at 37°C for 30 minutes.
During this 30 minute incubation, you may want to set up your tyrosine standard dilutions, which is done using 6 dram vials (dram vials can be substituted with polypropylene tubes) that can easily hold 8 mls. To the six vials the 1.1 mM tyrosine standard stock solutions is added with the following volumes in mls: 0.05, 0.10, 0.20, 0.40, 0.50. Don't add any tyrosine standard to the blank. Lower standards may be needed for impure test samples with that will yield little color change. Once the tyrosine standard solution has been added, add an appropriate volume of purified water to each of the standards to bring the volume to 2 mls.
After the 30 minute incubation, filter each of the test solutions and the blank using a 0.45 um polyethersulfone syringe filter. Filtration is required to remove any insolubles from the samples. The filtration 2 mls of the test samples and blank filtrate is then added to 4 dram vials that can hold at least 8 mls. You can use the same type of vial in which the standards were prepared. To all of the vials containing the standards and standard blank, 5mls of sodium carbonate is added, and for best results, 1 ml of Folin’s reagent is added immediately afterwards. Sodium carbonate is added to regulate any pH drop created by the addition of the Folin’s reagent. Sodium carbonate is then added to our test samples and test blank. You’ll notice that these solutions become cloudy after the addition of sodium carbonate. Then, the Folin’s reagent is added, which will react primarily with free tyrosine. The dram vials are then mixed by swirling and incubated at 37ºC for 30 minutes.
After this incubation, you should notice that the standards have a gradation of color correlating with the amount of tyrosine added; the highest concentrations of tyrosine appearing darkest. You can also notice appreciable color change in our test samples. 2mls of these solutions are filtered using a 0.45 um polyethersulfone syringe filter into suitable cuvettes. Now we performed the assay, we can proceed to the spectrophotometer to record our absorbance values.
Measuring Absorbance and Calculating Enzyme Activity
The absorbance of our samples is measured by a spectrophotometer using a wavelength of 660nm. The light path is set to 1cm. Record the absorbance values for the standards, standard blank, the different test samples, and test blank. Once all of the data has been collected, we are ready to create our standard curve. In order to generate the curve, difference in absorbance between the standard and standard blank must be calculated. This is the absorbance value attributable to the amount of tyrosine in the standard solutions. After this simple calculation, we create our standard curve using a graphing program by plotting the change in absorbance of our standards on the Y axis, versus the amount in micromoles for each of our 5 standards on the X axis.
| Volume of Tyrosine Standard | uMoles Tyrosine | |
| 0.05 | 0.055 | |
| 0.10 | 0.111 | |
| 0.20 | 0.221 | |
| 0.40 | 0.442 | |
| 0.50 | 0.553 | |
Once we have entered in our data points, generate a line of best fit and corresponding slope equation.
We then find the change in absorbance in our test samples by calculating the difference between our test sample absorbance and the absorbance of our test blank. Inserting the absorbance value for one of the test samples into the slope equation and solving will result in the micromoles of tyrosine liberated during this particular proteolytic reaction. To get the activity of enzyme in units per/ml, perform the following calculation.
Units/ml Enzyme = | (umole tyrosine equivalents released) x (11) | |
11= Total volume (in milliliters) of assay
10= Time of assay (in minutes) as per the Unit definition
1= Volume of Enzyme (in milliliters) of enzyme used
2= Volume (in milliliters) used in Colorimetric Determination
Take the number of micromoles tyrosine equivalents released obtained from the slope equation and multiply it by the total volume of the assay in mls, which in our case is 11mls. Then, divide this value by three other quantities: the time of the assay, which we ran for 10 minutes, the volume of enzyme used in the assay, which was varied, - let's use 1ml - the volume of milliliters used in colorimetric detection, which may differ based on your cuvette. We used 2 mls.
Micromoles of tyrosine divided by time in minutes gives us our measurement of protease activity that we call units. We can cancel out the units for volume measurement in the numerator and denominator, and are hence left with a measurment of enzyme activity in terms of units/ml. In order to determine the activity in a solid protease sample diluted in enzyme diluent we divide our activity in units/ml by the concentration of solid used in this assay originally in mg/ml. Leaving us with activity in terms of units/mg.
Units/mg solid = | Units/ml enzyme | |
Observation:
Results and Discussion:
Conclusion:
We've just shown you how to analyze protease activity of crude extract. In addition, this assay is useful to ensure that our proteases have precisely determined activity before you receive them for your experiments. As you have seen, when doing this procedure it's paramount to remember to heat both the casein and tyrosine solutions slowly and not to boil them as boiling will cause degradation of the protein and effect results of the assay. Also, it's critical to prepare different blanks for both your standards and for each test sample that you have.
References:
1. Anson, M.L., (1938) J. Gen. Physiol. 22, 79-89
2. Folin, O. and Ciocalteau, V., (1929) J. Biol. Chem. 73, 627
3. Proteolytic enzymes: A pratical approach. Beynom,R.J., Bond, J.S. (eds).1989. Academic press. Oxford.
4. Rawling, N.D., Barret, A. 1994. Families of serine peptidases. Meth. Enzymol. 244:18-61
5. Rawling, N.D., Barret, A. 1994. Families of cysteine peptidases. Meth. Enzymol. 244:461-486.
6. Rawling, N.D., Barret, A. 1994. Families of aspartic peptidases, and those of unknown mechanism. Meth. Enzymol. 248:105-120.
7. Rawling, N.D., Barret, A. 1994. Families of metallopeptidases. Meth. Enzymol. 248:183-228.
8. Rao,M.B., Tanksale, A .P., Ghatge, M.S., Deshpande,V.V.1998. Molecular and biotechnological aspects of microbial proteases, microbiology and molecular biology reviews,62 (3):597-635.
9. Alvarez-sanchez, M.E.A., Avila-Gonzalez,L., Becerril-Garcia, C., Fattel-Facenda,L.F., Ortega -Lopes, J. & Arroyo, R.2000. A novel cysteine proteinase (CP65) of Trichomonas vaginalis involved in citotoxicity. Microbial pathogenesis, 28:193-202.
10. http://www.wikipedia.com