

Sunday, December 24, 2017
Wednesday, December 13, 2017

Bioterrorism; An emerging threat to World
Bioterrorism
A bioterrorism attack is
the deliberate release of viruses, bacteria, or other germs (agents) used to
cause illness or death in people, animals, or plants. These
agents are typically found in nature, but it is possible that they could be
changed to increase their ability to cause disease, make them resistant to
current medicines, or to increase their ability to be spread into the
environment. Biological agents can be spread through the air, through water, or
in food. Terrorists may use biological
agents because they can be extremely difficult to detect and do not cause
illness for several hours to several days. Some bioterrorism agents, like the smallpox
virus, can be spread from person to person and some, like anthrax, cannot.
History
— In
1984, in The Dalles, Oregon, U.S., a group of extremist followers of Bhagwan
Shree Rajneesh (also known as Osho) contaminated the salad in 10 different
salad bars with the pathogen of salmonellosis, Salmonella thyphimurium,
in order to disable the population. A
total of 751 people contracted the disease and several of them were
hospitalized. Although there were no fatalities, this terrorist act is
considered the largest bioterrorist attack in the history of the U.S. (Török et
al., 1997).
— In
the 1990s, the Japanese cult of Aum Shinrikyo tested different bioweapons,
including botulin toxin, anthrax, cholera, and Q fever.
— In
1993, during a humanitarian mission in Africa, it tried to obtain samples of
the Ebola virus.
— Between
1990 and 1995, the cult attempted to carry out several bioterrorist acts in
Tokyo using vaporized biological agents, including botulinum toxin and anthrax
spores. Fortunately, the attacks were unsuccessful (Olson, 1999).
— A
significant bioterrorist event occurred in the U.S. contextually to the
dramatic attacks to the World Trade Center in New York in September 2001. The
release of Bacillus anthracis spores through the U.S. postal system was
carried out with letters addressed to the press and to government officials. There
were 22 confirmed cases of anthrax contamination, consisting of 12 cutaneous
and 10 inhalational cases. The 12 cutaneous patients responded positively to
antibiotic treatment, while of the 10 inhalational cases, 4 were fatal
(McCarthy, 2001).
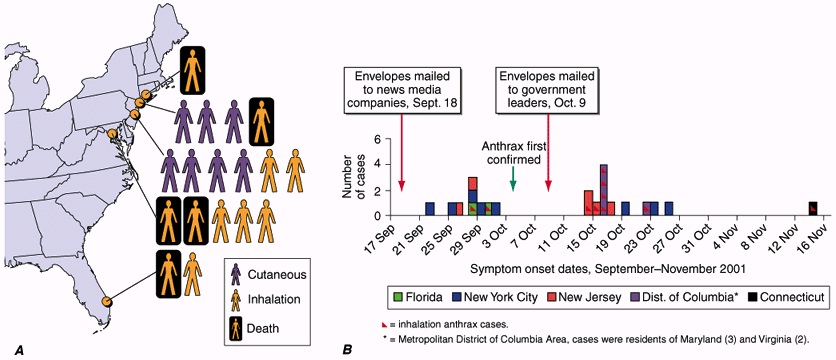
Figure showing Confirmed anthrax cases associated with bioterrorism: U.S.,
2001.
A. Geographic location and clinical manifestation of the 11 cases of confirmed inhalational and 7 cases of confirmed cutaneous anthrax.
B. Epidemic curve for the 18 confirmed cases of inhalational and cutaneous anthrax and 4 cases of suspected cutaneous anthrax.
A. Geographic location and clinical manifestation of the 11 cases of confirmed inhalational and 7 cases of confirmed cutaneous anthrax.
B. Epidemic curve for the 18 confirmed cases of inhalational and cutaneous anthrax and 4 cases of suspected cutaneous anthrax.
— In
2002, in Manchester, U.K., six terrorists were arrested for being found in
possession of ricin, and in 2004, traces of the same toxin were found at the
Dirksen Senate Office Building in Washington D.C. (Bhalla & Warheit, 2004).
— It
appears evident then that the use of biological agents has moved, in recent
times, to terrorist groups.
— This
creates very strong concerns that the use of bioweapons by terrorists can
create unexpected scenarios characterized by massive destructive potential
Bioterrorism agents’
important features of a perfect BW are:
- Highly
infectious and highly effective.
- Easily
produced with a long shelf life.
- Efficiently
dispersible.
- Readily
grown and produced in large quantities.
- Stable
on storage.
- Resistant
enough to environmental conditions.
- Resistant
to treatment
- High morbidity and mortality
- Potential for person-to-person spread
- Low infective dose and highly infectious by aerosol
- Lack of rapid diagnostic capability
- Lack of universally available effective vaccine
- Potential to cause anxiety
- Availability of pathogen and feasibility of
production
- Database of prior research and development
- Potential to be “weaponized”
Category of Bioterrorism
by Centers for Disease Control and Prevention (CDC):
The U.S. Centers for
Disease Control and Prevention (CDC) defines a bioterrorism attack as “the
deliberate release of viruses, bacteria or other germs (agents) used to cause
illness or death in people, animals, or plants” (CDC, 2013). It classifies biological agents into three
categories
Category A:
·
The U.S. public health system and primary
healthcare providers must be prepared to address various biological agents,
including pathogens that are rarely seen in the United States.
·
High-priority agents include organisms
that pose a risk to national security because they can be easily disseminated
or transmitted from person to person;
·
result in high mortality rates and have
the potential for major public health impact;
·
might cause public panic and social
disruption; and
·
require special action for public health
preparedness.
Groups
|
Diseases
|
Agents
|
A
|
Anthrax
|
Bacillus anthracis
|
Botulism
|
Clostridium botulinum toxin
|
|
Plague
|
Yersinia pestis
|
|
Smallpox
|
Variola major
|
|
Tularemia
|
Francisella tularensis
|
|
Viral hemorrhagic fevers
|
Filoviruses (e.g.
Ebola, Marburg) and Arenaviruses (e.g. Lassa, Machupo)
|
Category B:
— Second
highest priority agents include those that are moderately easy to disseminate;
— result
in moderate morbidity rates and low mortality rates; and
— require
specific enhancements of CDC's diagnostic capacity and enhanced disease
surveillance.
Groups
|
Diseases
|
Agents
|
B
|
Brucellosis
Epsilon toxin
|
Brucella spp.
Clostridium perfringens
|
Food safety threats
|
Salmonella spp.,
E.coli O157:H7, Shigella
|
|
Glanders
|
Burkholderia mallei
|
|
Melioidosis
|
Burkholderia pseudomallei
|
|
Psittacosis
|
Chlamydia psittaci
|
|
Q fever
|
Coxiella burnetii
|
|
Ricin toxin
|
Ricinus communis
|
|
Staphylococcal enterotoxin
B
|
Staphylococcus spp.
|
|
Typhus fever
|
Rickettsia prowazekii
|
|
Viral encephalitis
|
Alphaviruses (e.g.
Venezuelan equine encephalitis, Eastern equine encephalitis, Western equine
encephalitis
|
|
Water safety threats
|
Vibrio cholerae,
Cryptosporidium parvum
|
Category C:
— Third
highest priority agents include emerging pathogens that could be engineered for
mass dissemination in the future because of availability;
— ease
of production and dissemination; and
— potential
for high morbidity and mortality rates and major health impact.
|
Groups
|
Diseases
|
Agents
|
|
C
|
Emerging infectious diseases
|
Nipahvirus and
Hantavirus
|
Other classifications:
Generally, biological
agents (included those used as bioweapons) can be further classified according
to certain characteristics that define the hazard to health (NATO, 1996):
a.
Infectivity: The
aptitude of an agent to penetrate and multiply in the host.
b.
Pathogenicity: The
ability of the agent to cause a disease after penetrating into the body.
c.
Transmissibility: The
ability of the agent to be transmitted from an infected individual to a healthy
one
d.
Ability to neutralise: Its
means to have preventive tools and / or therapeutic purposes.
Transmissions:
Biological agents can be
transmitted through one or more ways.
The transmission modes are the following:
- Parenteral: Agents
that are transmitted through body fluids or blood.
- Airway (by droplets): Agents
that are emitted by infected people, which can then be inhaled by
surrounding people.
- Contact: Through
which the agents present on the surface of the infected organism can
infect another organism.
- Oral-faecal route: Through
objects, foods or other items contaminated with the faeces of infected
patients, or through sexual contact.
Impacts of Bioterrorism:
— Economic
impact of a bioterrorism attack could be devastating.
— Cost
$23 million to decontaminate a government building after 2001 anthrax attacks
in the US.
— Early
intervention can significantly decrease the costs resulting from a bioterrorist
attack.
— Still
expensive to provide prophylactic antibodies to a large number of individuals
— Reduction
in hospital admissions greatly outweighs initial costs
Warning signs:
— In
any location hit by a bioterrorism act the public health system will probably
be first to detect and respond.
— May
not be realistic to wait for confirmation of diagnosis.
— Delay
increases the potential for spread.
— Emergency
response may need to be activated on basis of patterns and timing of patient
presentation.
— Important
clues that can help alert hospitals to bioterrorist attack
— Every
health care professional should be suspicious of any unusual activity.
— It
will take many people in a variety of fields to control the impact of a
biological attack.
— Veterinarians
–many infectious diseases are zoonotic
— Scientists,
epidemiologists, doctors, and nurses will need to work together.
— Law
enforcement –reporting disease and controlling public reaction
— Bioterrorism
is a matter of national and international security.
— Require
the coordination of local, state, federal, and international agencies
Individual role
— It
is imperative that you understand your role.
— Prepare
ahead of time.
— Become
familiar with the location of important telephone numbers and resources.
— Then
you will be ready to assist at a moment’s notice.
— Your
day-to-day responsibilities may be much different during the response to a
bioterrorist attack.
— First
step is notifying the proper officials.
— Know
how to contact these agencies in advance.
— This
may save crucial minutes during a time of chaos.
Monday, November 20, 2017
DNA-THE DEOXYRIBONUCLEIC ACID
THE STRUCTURE AND FUNCTION OF DNA
Biologists in the 1940s had difficulty in conceiving how DNA could be the genetic
material because of the apparent simplicity of its chemistry. DNA was known to
be a long polymer composed of only four types of subunits, which resemble one
another chemically. Early in the 1950s, DNA was examined by x-ray diffraction
analysis, a technique for determining the three-dimensional atomic structure of
a molecule. The early x-ray diffraction results indicated that DNA was composed
of two strands of the polymer wound into a helix. The observation that DNA was
double-stranded was of crucial significance and provided one of the major clues
that led to the Watson - Crick Model for DNA structure. But only when this
model was proposed in 1953 did DNAs potential for replication and information
encoding become apparent. In this section we examine the structure of the DNA
molecule and explain in general terms how it is able to store hereditary
information.
HISTORY OF DNA EVOLUTION
·
In 1868: F. Miescher isolated nucleic acids from white
blood cells that were acidic in nature to which he called nuclein.
·
In 1880: Fischer isolated purines and pyrimidines.
·
In 1881: Zacharis identified nuclein with chromatin.
·
In 1899: Altaman replaced the term nuclein with nucleic
acid.
·
In 1900s: Kossel identified the presence of histones and
protamines with nucleic acids (Nobel laurate).
·
In 1910s: P. A. Levene discovered phosphate and pentose
sugars callled deoxyribose molecule.
·
In1928: Frederick Griffith demonstrate the existence of a
chemical in bacteria that caries genetic information
·
In 1943: Three American Microbiologist; Ostawald Avery,
Colin MacLeod and Maclyn McCarty for the first time presented the evidence that
DNA is the genetic material and is made up of genes.
·
In 1944: Oswald Avery showed that degradation of DNA and
not protein resulted in loss of genetic information.
·
In 1950s: Rosalind Franklin and her supervisor Maurice
Wilkins were working on the X-ray diffraction model for DNA.
Rosalind Franklin (1951):
·
Generated X-ray
crystallography data suggesting a double helix with phosphates on the outside
- Rosalind Franklin who actually proposed the concept of double helix was deprived of
Nobel prize due to cruel death in 1958.
- Great revolution in DNA Biology
- In 1953 February: Pauling and R. B. Corey gave a
triple helix model of the DNA molecule. However he couldn’t explain the
process of DNA replication.
- They were near to present about the double helix
model.
- In 1953 April: J.D. Watson (an American Biologist)
and F. H. C. Crick (a British Physicist) presented the double helix model
of DNA (published in Nature entitled “A structure for deoxyribose
nucleic acid’).
- Nobel prize awarded to
o
Watson, Crick and Wilkins
o
in 1962.
DNA STRUCTURE
DNA is composed of nitrogen bases, deoxyribose sugars and phosphate. Adenine
and guanine are purine bases while cytosine and thymine are pyrimidine bases.
The phosphodiester bond between sugar and phosphate molecules form the backbone
of DNA. The glycosidic bond is formed between nitrogen bases and sugar
molecules.

Figure 1: Nitrogen bases

Figure 2: Structure of nucleotide showing
phosphodiester bond and glycosidic bond

Figure 3: Hydrogen bonding between nitrogen bases
CHARGAFF EQUIVALENT RULE
In 1948, a chemist Erwin Chargaff, on the basis paper chromatography
experiment, analyze the base composition of DNA. In 1950, he discovered that: In
a DNA molecule of different types of organisms, the total no. of purines is
equal to the total no. of pyrimidines. A/T=G/C
Number of Purines (A+G) = Number of Pyrimidines (C+T)
WATSON AND CRICK MODEL FOR DNA
The was proposed by Watson and Crick which was published in Nature
in 1953. It is also known as ‘double helix model for DNA’
molecules. However, the photograph for model was taken from X-ray diffraction
photograph from Rosalind Franklin.

Figure 4: Double helix structure of DNA
According to the Model:
·
DNA molecule consists of two strands which are connected
by H-bonding and they are helically twisted.
·
Each step in one strand consists of nucleotide of purine
base which alternately pair with pyrimidine base.
·
DNA is a polymer of four nucleotides (A T G C).
·
Adenine pairs to thymine with 2-H bonding (A=T).
·
Gaunine pairs to cytosine with 3 H-bodings (GºC).
·
Two strands apart 20 A from each other.
·
Helix coils in right hand i.e. clockwise direction and
completes at every 34 A distance.
·
Two strands are complementary to each other.
·
One strand runs 5’®3’ while the complementary strand runs 3’®5’.
·
The polarity of DNA is due to direction of phosphodiester
linkage.
·
Turning results in deep and wide major groove which
is the site of bonding of specific protein.
·
The distance between two strands form a minor groove.
·
One turn of double helix at every 34 A distance includes 10
nucleotides.
·
Each nucleotide is situated at a distance of 3.4 A.
·
Sugar phosphate makes the back bone of double
helix of DNA molecules.
·
The DNA model also suggested a copying mechanism of the
genetic material which is semi conservative in nature.
·
Experimentally proved by Mathew, Meselson and Frank W.
Stahl in 1958.
·
Universally accepted.
DIFFERENT FORMS OF DNA
Three different forms of DNA are
found i.e. A form, B form and Z form. The B form (10 bp/turn), which is
observed at high humidity, most closely corresponds to the average structure of
DNA under physiological conditions. A form (11 bp/turn), which observed under
the condition of low humidity, presents in certain DNA/protein complexes. RNA
double helix adopts a similar conformation.
Z form (12 bp/turn) more loosely arranged DNA is found during DNA replication.

Figure 5: Different forms of DNA

A DNA Molecule Consists of Two Complementary Chains of Nucleotides:
A deoxyribonucleic acid (DNA) molecule consists of two long polynucleotide chains
composed of four types of nucleotide subunits. Each of these chains is known as
a DNA chain, or a DNA strand. Hydrogen bonds between the base portions of the
nucleotides hold the two chains together. The nucleotides are composed of a
five-carbon sugar to which is attached one or more phosphate groups and a
nitrogen-containing base. In the case of the nucleotides in DNA, the sugar is
deoxyribose attached to a single phosphate group (hence the name
deoxyribonucleic acid), and the base maybe either adenine (A), cytosine (C),
guanine (G), or thymine (T). The nucleotides are covalently linked together in
a chain through the sugars and phosphates, which thus form a
"backbone" of alternating sugar-phosphate. Because only the base
differs in each of the four types of subunits, each polynucleotide chain in DNA
is analogous to a necklace (the backbone) strung with four types of beads (the
four bases A, C, G, and T). These same symbols (A, C, G, and T) are also
commonly used to denote the four different nucleotides-that is, the bases with
their attached sugar and phosphate groups. The way in which the nucleotide
subunits are linked together gives a DNA strand a chemical polarity. If we
think of each sugar as a block with a protruding knob (the 5'phosphate) on one
side and a hole (the 3'hydroxyl) on the other, each completed chain, formed by
interlocking knobs with holes, will have all of its subunits lined up in the
same orientation. Moreover, the two ends of the chain will be easily
distinguishable, as one has a hole (the 3'hydroxyl) and the other a knob (the
5'phosphate) at its terminus. This polarity in a DNA chain is indicated by
referring to one end as 3' end and the other as the 5' end. The
three-dimensional structure of DNA-the double helix-arises from the chemical
and structural features of its two polynucleotide chains. Because these two
chains are held together by hydrogen bonding between the bases on the different
strands, all the bases are on the inside of the double helix, and the sugar-phosphate
backbones are on the outside. In each case, a bulkier two-ring base (a purine)
is paired with a single-ring base (a pyrimidine); A always pairs with T and G
with C.
DNA TOPOLOGY
In order to fully
understand DNA topology, students need to familiarize themselves with three key
mathematical concepts: twist (Tw), writhe (Wr), and linking number (Lk). Twist
represents the total number of double helical turns in a given segment of DNA.
By convention, the right-handed twist of the Watson-Crick structure is assigned
a positive value. Writhe is a property of the spatial course of the DNA and is
defined as the number of times the double helix crosses itself if the molecule
is projected in two dimensions. The helix-helix crossovers (i.e., nodes)
are assigned a positive or negative value based on the orientation (i.e.,
handedness) of the DNA axis. The numerical term that describes the sum of the
twist and the writhe is called the linking number, which represents the total
linking within a DNA molecule. Mathematically, these properties of DNA can be
expressed as:
Lk = Tw + Wr
Why is DNA supercoiling important?
Duplex DNA is merely the storage form for the genetic information. In order to
replicate or express this information, the two strands of DNA must be
separated. Since the global underwinding of the genome imparts increased
single-stranded character to the double helix, negative supercoiling greatly
facilitates this process. As a result, replication origins and gene promoters
are more easily opened, and rates of DNA replication and transcription are
greatly enhanced.
While negative supercoiling
promotes many DNA processes, positive supercoiling inhibits them. When tracking
systems, such as replication or transcription complexes travel along the double
helix, they do not spiral circumferentially around the DNA. Rather, they move
linearly through the DNA and the double helix spins to accommodate this motion.
Recall from the earlier discussion that the ends of chromosomal DNA are not
free to rotate. As a result, the number of turns of the helix remains invariant
unless the nucleic acid chain is broken. Thus, the linear movement of tracking
enzymes through DNA does not change the number of turns, but merely compresses
them into a shorter segment of the genetic material. Consequently, the double
helix becomes increasingly overwound ahead of tracking systems. DNA
overwinding, or positive supercoiling, makes it more difficult to open the two
strands of the double helix and ultimately blocks essential nucleic acid
processes if not alleviated.

Figure 6: DNA supercoiling
Topoisomerases I
Type I topoisomerases are denoted by “odd” numbers (topoisomerase I,
III, etc.). These enzymes are monomeric in nature and require no high-energy
cofactor. There are two subclasses of type I enzymes, type IA and type IB. Type
I topoisomerases act by creating transient single-stranded breaks in the double
helix, followed by passage of the opposite intact strand through the break
(type IA) or by controlled rotation of the helix around the break (type IB).
Type IA enzymes require divalent metal ions for catalytic activity and
covalently attach to the 5’-terminal phosphate of the DNA. In contrast, type IB
enzymes do not require divalent metal ions and covalently attach to the
3’-terminal phosphate
Topoisomerases II
Type
II topoisomerases are denoted by “even” numbers (topoisomerase II, IV, etc.).
These enzymes contain multiple polypeptide chains and require ATP for overall
catalytic activity. Prokaryotic enzymes have an A2B2 structure and eukaryotic enzymes are
homodimers in which the bacterial A and B subunits have merged. Based on the
structure of the archetypical bacterial type II enzyme, gyrase (see below), the
A subunit (or domain) contains the active site tyrosyl residue that links to
DNA during the cleavage event and the B subunit (or domain) contains the site
of ATP hydrolysis.
Type II topoisomerases modulate DNA
topology by generating a transient double-stranded break in the DNA backbone,
passing a separate double helix through the opening, and resealing the break.
All bacterial and eukaryotic type II enzymes require divalent metal ions for
activity and those examined so far appear to utilize a two-metal-ion mechanism
similar to that of DNA polymerases and primases. The cleavage reaction of type
II topoisomerases generates DNA intermediates with 4-base, 5’-cohesive ends
that are covalently attached to the enzyme through their 5’-terminal
phosphates
Bacteria in Photos





















